Clupeomorpha fishes are divided into two main groups: Clupeoformes (such as sardines and Yangtze knife fish) and the extinct Ellimmichthyiformes. Ellimmichthyiformes, commonly known as double-armored herrings, were first recorded in the Early Cretaceous and continued into the Oligocene (Oligocene material is still under study). They were once widely distributed in South America, West Africa, North America, the Mediterranean coast, and East Asia. Most known species originated in marginal marine environments, while those from the Cretaceous onwards originated in freshwater environments influenced by marine environments, such as the Eocene Green River Shale in North America and the Shengli Oilfield in East China, as well as southern Guangxi in the Oligocene. Among these, the Shengli Double-armored Herring (Diplomystus shengliensis) from the Shengli Oilfield, due to its high similarity to different species of the same genus in the contemporary Green River Shale in North America, is one of the most important pieces of evidence for the "trans-Pacific distribution pattern" of Eocene fish fauna.
Recently, the *Journal of Vertebrate Paleontology* published online the research findings led by Fei-Xiang Wu's team at our institute, with the participation of geologists from Carthage University and El Manar University in Tunisia. Based on new material discovered in the Lower Cretaceous strata of the Chotts Basin in southern Tunisia, a new genus and species of herring (*Tunisiaclupea speratus*) was established within the family Paracoilidae of the order Eleocharis. This discovery supplements the early record of this group in the Mediterranean region (Western Tethys Ocean) and provides new information for studying the phylogenetic relationships and biogeographical history of *Tunisiaclupea* (Figures 1-3).
The Ellimma branneri currently comprises 13 genera and at least 35 valid species. Since the establishment of this order (Grande, 1985), previous studies have yielded many discrepancies regarding their internal taxonomic relationships and biogeographical history. This study, based on new fossil material and a phylogenetic matrix containing the most complete range of species in this order (Figure 3), classifies the Tunisian herring into the Paraclupeidae family (Chang and Chou, 1977) and adjusts the internal taxonomic relationships within the Paraclupeidae family. As a relatively primitive type within the Paraclupeidae family, the Tunisian herring phylogenetically lies between the Early Cretaceous *Ellimma branneri* of Brazil and the Late Cretaceous *Tycheroichthys dunveganensis* of northern North America.
The Mediterranean region is a key area in the evolutionary history of the Eglispidae. Some major lineages of this order (such as Armigatidae and Sorbinichthyidae) may have originated in this region during the Late Cretaceous. Meanwhile, the Mediterranean, as an important block in the evolution of the Tethys Ocean, has undergone extensive marine-terrestrial changes since the Cretaceous, with continuous changes in its aquatic environment. Tunisia, located on the southern edge of the Western Tethys Ocean during the Cretaceous, was situated at the opening of a waterway connecting the Tethys Ocean and the expanding South Atlantic Ocean, which was another hotspot in the evolutionary history of the Eglispidae. Previously, the fossil record of Eglispidae in Tunisia was scarce, with only the Late Cretaceous *Diplomystus solignaci*. In recent years, Carthage University and El Manar University of Tunisia have conducted joint excavations in the Chotts Basin in southern Tunisia, finding abundant and well-preserved aquatic fossils in the Lower Cretaceous strata (Figure 1). The newly discovered Tunisiaclupea speratus is the earliest fossil record of the Hermitae order in Tunisia and even on the southern edge of the Western Tethys Ocean.
This study, based on new phylogenetic relationships, reconstructs the zoogeographical history of the *Eleocharis eleocharis* and offers a new explanation for its diversity development. We believe that the diversity of *Eleocharis eleocharis* in the Western Tethys Ocean (present-day Mediterranean) reached its peak during the Late Cretaceous Cenomanian period, possibly related to the high sea levels and widespread marine transgression at that time. When the Western Tethys Ocean connected with the tropical Atlantic Ocean from a surface sea running north of Africa, habitat expansion, the influx of warm currents, and increased food input promoted their migration and dispersal, leading to enhanced allophagocytic speciation. This group gradually declined after the Cretaceous, with the youngest current record coming from Ningming, Guangxi, during the Oligocene.
Based on current understanding, the earliest representatives of the Elim herring were distributed in South America, the Mediterranean, and East Asia (Paraclupea, an Early Cretaceous herring from Zhejiang). The biogeographical connection between the former two has been supported by phylogenetic evidence. However, the vast area between the Mediterranean and East Asia, including West Asia, South Asia, and even southern Tibet—once part of the Neo-Tethys Ocean—has remained a blank area in its fossil record. As the new fish's name, "Hopeful Tunisian Herring," suggests, it is hoped that with the advancement of future research, the final piece of this "puzzle" will eventually be completed.
This research was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences, the Center for Excellence in Biological Evolution and Environment, the National Natural Science Foundation of China, the Tunisian Ministry of Scientific Research Fund, and the Youth Innovation Promotion Association of the Chinese Academy of Sciences.
Original link:
https://www.tandfonline.com/doi/full/10.1080/02724634.2018.1529675
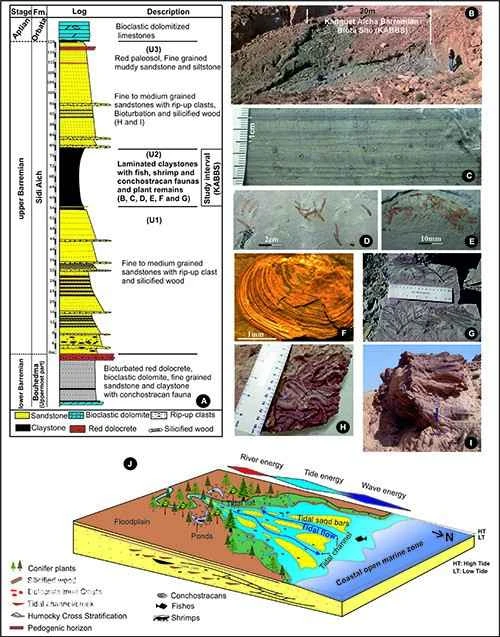
Figure 1. Fossil-bearing strata (AC), fossil-bearing deposits (DI), and paleoenvironmental reconstruction (J) of the Chotts Basin in Tunisia. Image courtesy of Wu Feixiang.
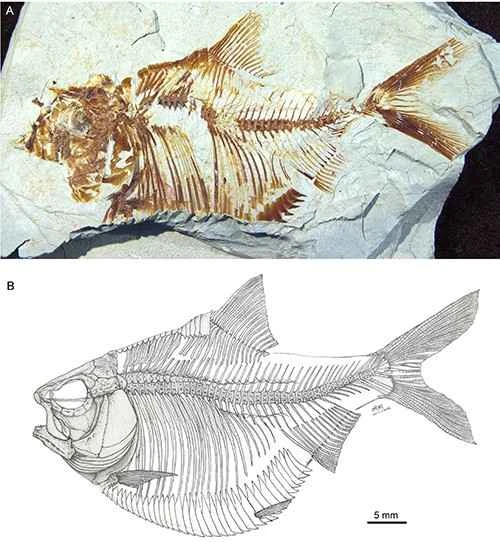
Figure 2. Homotype of the Tunisian herring (A) and skeletal reconstruction (B). Photo provided by Wu Feixiang.
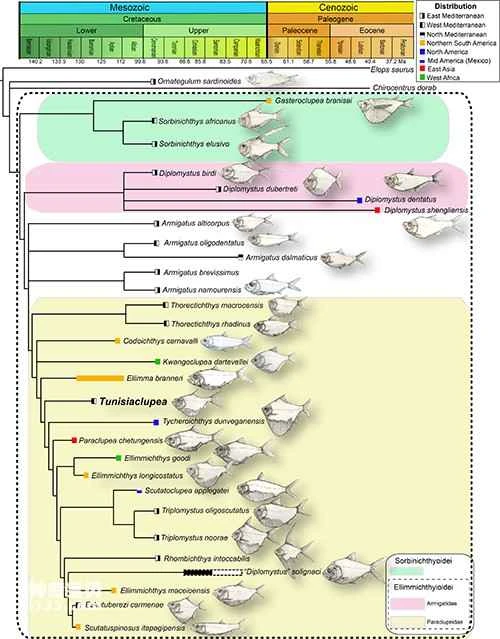
Figure 3. Phylogenetic analysis results. Image provided by: Wu Feixiang


